
CÁNCER DE COLON
Mundialmente el cáncer de colon (CCR) ocupa el tercer lugar en incidencia y el cuarto en mortalidad a nivel mundial. Según estadísticas de E.U.A, la tasa de incidencia es de 46/100,000, una media de edad de aparición de 69 años, sobrevida a 5 años de 64% y un riesgo acumulado de 5% (es decir, uno de cada veinte estadounidenses lo padecerá a lo largo de su vida). El Instituto Nacional de Estadística y Geografía (INEGI) reportó 74,685 muertes por cáncer en 2010 (13% de las muertes en México) de las cuales 5.4% son por CCR.
Las manifestaciones clínicas del CCR se relacionan con el tamaño y la localización de la tumoración. Entre los signos y síntomas frecuentes de las neoplasias proximales (ciego y ángulo esplénico) existe el dolor abdominal indefinido, pérdida de peso corporal importante y sangrado oculto en eses. En cambio, las neoplasias distales (colon descendente a recto) pueden presentarse con alteraciones en los hábitos intestinales, disminución del calibre de la deposición, y en algunos casos está presente la hematoquezia.
Dentro de la fisiopatología es importante conocer los elementos que la patogénesis molecular ha tomado en los últimos años, quedando más que demostrado que la progresión morfológica de la mucosa normal a adenoma, y de éste a tumor maligno, es debido a mutaciones del ADN. Para que el CCR aparezca se necesita una serie de mutaciones acumuladas en genes específicos conocidos como genes supresores tumorales, oncogenes y genes reparadores del ADN.
Para poder entender esto de una manera más sencilla, es importante conocer los conceptos de cada uno de estos genes; es así que, los genes supresores tumorales son aquellos que normalmente suprimen y regulan el crecimiento celular; para inactivarlos, ambas copias del gen deben mutar o perderse; cuando esto ocurre se disminuye en algún grado la regulación del crecimiento. Por otro lado, los oncogenes estimulan el crecimiento celular y cuando éstos mutan, la estimulación del crecimiento se encuentra continuamente activada. La mutación de una sola copia de oncogen es suficiente para que ocurra este cambio. Mientras tanto, la función normal de los genes reparadores del ADN es arreglar las mutaciones que ocurren durante la división celular o como una consecuencia de eventos que dañan el ADN. Cuando estos genes funcionan de manera inadecuada persisten mutaciones en muchos otros afectando importantes vías moleculares que conducen a la progresión del cáncer como se ejemplifica en la imagen siguiente.

VÍAS EN EL DESARROLLO DE CÁNCER DE COLON
Los estudios moleculares de la CCR han dilucidado que varias vías de señalización están involucradas en su desarrollo. Ahora tenemos una idea bastante detallada de los principales eventos genéticos involucrados en la carcinogénesis colorrectal. La heterogeneidad morfológica, en términos de sitio, grado y tipo del tumor, se ha ampliado por complejidad molecular , identificándose vías moleculares que actúan en el desarrollo de la neoplasia, como se observa en la imagen siguiente.
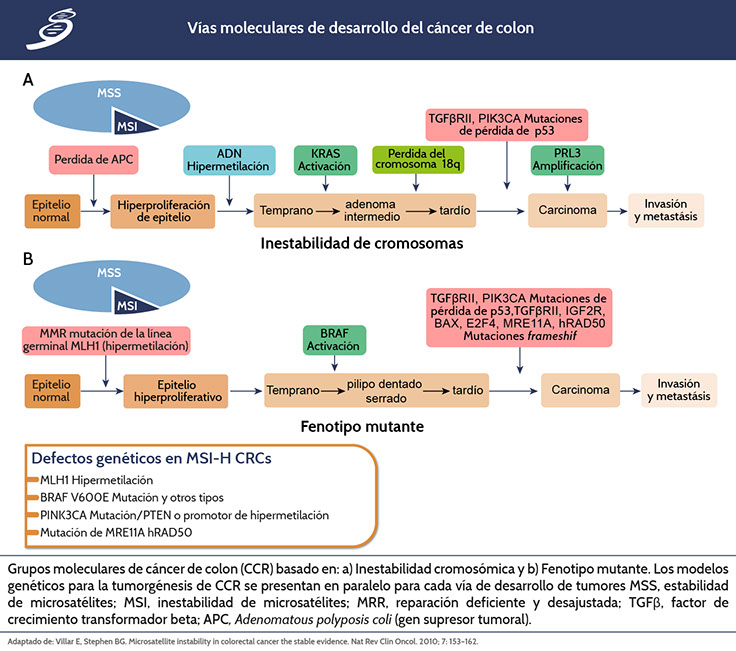
Vía clásica
De la cual la inestabilidad cromosómica (CIN) es un sello distintivo. Estos carcinomas suelen ser altamente aneuploides y muestran una sorprendente variación en el número de copias de genes; está involucrado en aproximadamente el 80% de los carcinomas colorrectales presentando anormalidades en los genes que a continuación se describen.
PAC: La mutación más temprana en la progresión al cáncer de colon es en el gen de la poliposis adenomatosa coli (PAC), un gen supresor tumoral, normalmente esta proteína se asocia con la betacatenina produciendo su destrucción. Cuando la PAC muta no puede unirse a la betacatenina permitiendo que los niveles intranucleares de ésta aumenten uniéndose al ADN, activando la trascripción de genes e inhibiendo la apoptosis que es responsable de la formación de adenomas. Una vez que el gen es inactivado en una célula única, la célula presenta una ventaja de crecimiento selectivo sobre las otras células progresando a un pólipo visible. Al menos 80% de los adenomas comienzan con una mutación del gen PAC como el primer paso en la patogénesis del cáncer de colon. Este gen es el responsable de la poliposis adenomatosa familiar al intervenir en la señalización Wnt, que desempeña un papel central por lo que cerca del 90% de todos los casos de CCR geneológicas se producen al activar esta vía.
Después de que el gen PAC pierde su función, mutan subsecuentemente muchos otros genes en el cáncer colorrectal como: K-ras, p53, DCC (Supresor del carcinoma colorrectal, por sus siglas en inglés) y los genes reparadores de la alteración en el acople del ADN, todos los cuales son supresores tumorales a excepción de K-ras, que es un oncogen. Ahora bien, K-ras está normalmente involucrado en la señal de transducción de crecimiento intracelular, por lo tanto, cuando una copia del K-ras muta, la señal de crecimiento es constante.
MMR: Los genes de reparación de falta de concordancia (MMR) son los encargados de reparar los errores en la replicación del ADN de uno o varios nucleótidos consecutivos que ocurren frecuentemente durante la división celular normal; prácticamente sirven como un sistema de revisión de errores moleculares. Hasta ahora existen seis genes identificados que participan en el proceso de reparo que son: hMLH1, hMSH2, hMSH3, hMSH6, hPMS1 y hPMS2. Cuando uno de estos genes es inactivado por mutaciones en ambas copias da como resultando errores persistentes en la replicación del ADN.
p53: Cuando el ADN presenta mutaciones en cualquiera de los cromosomas, el gen p53 induce a que el ciclo celular G1 se detenga. Esta función da el tiempo suficiente para el reparo de la mutación, y si éste no se reparó, el p53 induce la muerte celular apoptótica. Sin embargo, cuando ambas copias p53 mutan, el gen no es funcional, las mutaciones a través del genoma persisten y se acumulan; las mutaciones de este gen están presentes en al menos el 50% del CCR además de las mutaciones del p53, las cuales son eventos tardíos; en los adenomas ocurren más tempranamente antes de que se reconozca la displasia histológica.
Vía de inestabilidad de los microsatélites (min)
Esta vía es responsable del desarrollo de carcinomas hipermutadores, que en contraste con los carcinomas CIN suelen mostrar poca aneuploidía. El prototipo de estas vías se descubrió a principios de los noventa en la búsqueda de los orígenes genéticos moleculares del cáncer de colon hereditario no poliposis, que ahora se conoce como síndrome de Lynch. Un descubrimiento histórico fue que el síndrome de Lynch es causado por una mutación en uno de los genes ya mencionados. Los que también actúan en esta vía son los MMR encargándose de la corrección de errores, tales como desajustes o supresiones de una sola base e inserciones cortas, éstas son secuencias repetitivas de ADN, en su mayoría 1-6 pares de bases de longitud que se producen a lo largo del genoma, conocidas también como microsatelites (de ahí el nombre de esta vía). En la actualidad, la deficiencia de MMR genera nuevas secuencias de microsatélites, que pueden detectarse mediante un ensayo basado en PCR. En éste, un panel de loci de los marcadores de microsatélites, incluyendo BAT25, BAT26, D2S123, D5346 y D17S250, se utiliza para detectar la inestabilidad de éstos (MSI). Cuando dos o más marcadores son inestables, se llaman MSI-H (igh); cuando sólo un marcador es inestable, se llama MSI-L (ow); y cuando todos los marcadores son estables, se llaman microsatélites estables (MSS). Dado que las MRR son fácilmente detectables en secciones de tejido rutinariamente procesadas, la detección inmunohistoquímica de la pérdida de expresión de una de las proteínas relevantes (MLH1, MSH2, MSH6 y PMS2) se ha convertido en corriente principal en la detección del síndrome de Lynch.
En términos de ruta, MSI-H es equivalente a la inestabilidad vía microsatélite (MIN), esto se debe a la metilación del promotor de MLH1, que cierra la transcripción MLH1, resultando en MMR deficiente y MSI-H. Los carcinomas MSI muestran una frecuencia relativamente alta de mutaciones del gen BRAF. Existen otros genes mutados preferentemente, a menudo caracterizados por tramos mononucleótidos en sus regiones codificantes, como es el caso del gen del receptor TGF-β II y del gen proapoptótico BAX, que son, en consecuencia, frecuentemente inactivados en el cáncer deficiente en MMR.
La ruta del fenotipo de la hipermetilación de la isla CpG
La hipermetilación de islas CpG asociadas al promotor de genes, implicadas en la regulación de la transcripción, es un mecanismo frecuentemente encontrado responsable de silenciar genes supresores de tumores. Si este fenómeno se produce en todo el genoma, se llama hipermetilación de islas CpG (CIMP). Lo que causa el fenotipo CIMP y cuáles son exactamente sus consecuencias sigue siendo difícil de alcanzar. Se ha obervado que las lesiones mucosas planas no adenomatosas, especialmente en el colon derecho, a menudo presentan CIMP; estas lesiones se producen como resultado de la retención anormal del epitelio superficial debido a hiperproliferación e inhibición de la pérdida de células apoptóticas. Se observó que los carcinomas con CIMP estaban asociados con lesiones precursoras dentadas, y además los carcinomas de CIMP a menudo albergan mutaciones de BRAF. Los eventos moleculares en la vía incluyen además de la mutación CIMP del gen BRAF, específicamente la mutación V600E, metilación del promotor de una variedad de genes (como resultado de CIMP) y posterior metilación del promotor del gen MLH1 con silenciamiento de la expresión de MLH1 como resultado. Esto es entonces responsable de la insuficiencia de MMR, MSI, y un estado hipermutador. Es importante enfatizar aquí que estas nociones de vía constituyen un marco modelo que es útil en la definición adicional de eventos moleculares en el cáncer colorrectal en asociación con parámetros clínicos y patológicos.
Actualmente, se están realizando esfuerzos para desarrollar una clasificación molecular de consenso en el contexto del análisis de datos genómicos colaborativos de código abierto, cuyos primeros resultados ya se han publicado. Queda por ver hasta qué punto estas clasificaciones moleculares tendrán un impacto en el manejo clínico del CCR, que todavía está ampliamente determinado por los parámetros TNM clásicos, a pesar de que recientemente el estatus MS ha ganado importancia como parámetro molecular a tener en cuenta en la elección de terapia adyuvante de la fase II / III y potencialmente también la elección del tratamiento del metastásico, aunque aún está por comprobarse del todo si es así como estos conceptos de la vía han influido en el manejo clínico de la CRC.
Referencias
1. Rosty C, Young JP, Walsh MD, et al. Colorectal carcinomas with KRAS mutation are associated with distinctivemorphological and molecular features. Mod Pathol. 2013;26:825-34.
2. Bosman F, Yan P. Molecular pathology of colorectal cancer. Pol J Pathol. 2014;65:257-66.
3. Parra JFC. ONSENSO NACIONAL CÁNCER COLORRECTAL DIAGNÓSTICO Y TRATAMIENTO. Revista venezolana de oncología [Junio 2012] Disponible en: Consultado en: http://www.redalyc.org/pdf/3756/375634872006.pdf. [Revisando en Agosto 2017]
4. American Cancer Society. Colorectal Cancer Facts & Figures 2017-2019 [2017] Disponible en: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/colorectal-cancer-facts-and-figures/colorectal-cancer-facts-and-figures-2017-2019.pdf. [Revisando en Agosto 2017]
5. Benson Al B. Colon cancer National. Comprehensive Cancer Network (NCCN). Clinical Practice Guidelines in Oncology. Versión 2.2016 [Noviembre 2012] Disponible en: https://www.tri-kobe.org/nccn/guideline/colorectal/english/colon.pdf. [Revisando en Agosto 2017]
6. Roche. Colorectal cancer. A guide for journalists on colorectal cancer and its treatment [2012] Disponible en: http://www.roche.com/dam/jcr:8e08ab40-4ae0-4c61-99dd-a139cce45edc/en/med-colorectal-cancer.pdf. [Revisando en Agosto 2017]
7. Colussi D, Brandi G, Bazzoli F, et al. Molecular Pathways Involved in Colorectal Cancer: Implications for Disease Behavior and Prevention. Int J Mol Sci. 2013; 14: 16365–16385.
8. Villar E, Stephen BG. Microsatellite instability in colorectal cancer the stable evidence. Nat Rev Clin Oncol. 2010; 7: 153–162
9. Walther A, Johnstone E, Swanton C, et al. Genetic prognostic and predictive markers in colorectal cancer. Nat Rev Cancer. 2009;9:489-99
10. Kumar V, Abul KA , Mitchell R, Basic Pathology: With STUDENT CONSULT Online Access. 8 ed, Estados Unidos, Saunders;2007
Periodismo científico cultural
Sobre el cigarro electrónico
Ya ha pasado tiempo desde que el cigarro se convirtió en el enemigo público número uno, tanto para fumadores como para no fumadores...
[leer mas]
la sal! Tu cerebro
y corazón lo agradecerán
La enfermedad diverticular y sus complicaciones son una causa importante de morbimortalidad en todo el mundo...
[leer mas]
El primer anticuerpo monoclonal para pacientes con hemofilia A e inhibidores a FVIII
Cuando se vive con hemofilia todo puede ser una amenaza para provocar sangrados; desde niños...
[leer mas]
Infografías




TODOS LOS DERECHOS RESERVADOS © SPG COMUNICACIONES SA DE CV 2012-2018
Diseño: A. Victoria Pérez
Aviso de privacidad